Thinking with space: How mental representations of space bolster cognitive processes
The concept of thinking with space may seem as ubiquitously obvious as the answer to the question posed by David Foster Wallace’s (2009) fictional fish: “what is water”? For, how would it be possible to do anything without space? Of course we think with space; we exist in space, neurons take up space, and all we see relies on an underlying spatial framework. However, not everything we think about is overtly spatial. For instance, I need not account for space when preparing my outfit for the day nor when calculating a budget for food expenses next month. Nonetheless, the conceptualization of space is arguably the most fundamental cognitive procedure our brains participate in.
In order to posit the claim that spatial processing is man’s greatest cognitive feat, it would be prudent to reason as to what evolutionary constraints would foster such a resultant system. If an organism it to achieve its primary objective (i.e. permanence of existence), then it must acquire the resources to persevere and avoid environmental stimuli associated with its demise. From an evolutionary standpoint, the rudimentary component to accomplish this objective is movement—the act of changing one’s physical location within a larger spatial construct. Although animals without a densely centralized nervous system (e.g. organisms in the phylum Cnidaira – jellyfish) move, it has been proposed that the development of a brain was a tool in the evolutionary pursuit of producing movement:
We have a brain for one reason and one reason only, and that’s to produce adaptable and complex movements. There is no other reason to have a brain. Think about it. Movement is the only way you have of affecting the world around you. Now that’s not quite true. There’s one other way, and that’s through sweating. But apart from that, everything else goes through contractions of muscles. (Wolpert, 2011)
The life cycle of a clabelina moluccensis, more commonly referred to as the bluebell tunicate or blue sea squirt, is perhaps the most literal encapsulation of this proposition; the bluebell tunicate begins to digest its cerebral ganglion[1] once identifying a suitable undersea perch to spend the rest of its existence, reverting to the more primitive, sessile adult form of the species (Linas, 2001). The nutrients provided to the organism by digesting its nervous system is more valuable in regards to its prolonged existence than the retention of an advanced neural framework. Both Dennett (1993) and Linas (2001) took this evidence to suppose that a brain is necessary only for actively moving creatures.
I will argue hereafter that since a representation of space is a prerequisite for an organism’s ability to move (perhaps the most potent evolutionary prowess) that the brain’s spatial processing framework is it’s most powerful feature. Seeing as navigation about a spatial environment was the first concern of organisms, it has had the “head-start” of greatest breadth when compared to any and all other aspects of cognition. I will provide examples which illustrate that this categorical advantage has warranted the brain’s recycling of cognitive systems supporting spatial representation for the utility of non-spatial processing. It is this reutilization that I refer to when using the phrase thinking with space. By expanding beyond the boundaries of its initial goal-states, the brain’s spatial processing framework has afforded temporal, lingual, logical, and mnemonic systems with an advanced toolkit for achieving their more advanced organism-specific evolutionary contributions.
The advent of calendars, dating back to Near-East civilizations over four thousand years ago (Brittanica –Babylonian Calendars, 2015), marks the first time that intelligent creatures conceptualized time within a spatial construct. Calendars allow, to this day, one to explicitly visualize time in an organized fashion. Ancient astronomers lacked the ability to see the moon rotate the Earth, or the Earth move around the sun. However, they could note the temporal periodicity of the moon’s appearance in the sky and transform this temporal representation into a spatial one that treated each day as a unit with units to the left and right of it indicating previous and subsequent days, respectively. Such a transformation allowed for ancient civilizations to achieve initial insights as to the definite recurrence of these temporal patterns. In so doing, they were able to reliably plan for cosmic events that aided in agriculture. For example, the Egyptian civilization used calendars to calculate agricultural seasons around the flooding of the Nile – Akhet, the season of the inundation (Kemp, 2006).
By transforming a temporal phenomenon into a spatial dimension by way of a formal calendar system, ancient civilizations were able to recruit their evolved neural systems for spatial processing so as to conceptualize a previously intangible reality. The consequences of such an understanding allowed for a greater guaranteed chance of nutrition for the culture, which, ultimately led to longer lives and healthier offspring capable of fostering more advanced brains in the future. Furthermore, the opportunities afforded by the civic adoption of the calendar[2] are ubiquitous. By having a calendar at one’s fingertips, anyone can strategize in a coordinated fashion so as to organize for advanced sociological proceedings in a collaborative fashion. Such initially organized events like market fairs, town forums, and ovulation cycles led to the temporally coordinated modern events like music festivals, invasions, vote-days, and rocket launches. It’s difficult to imagine such modern civic feats of synchronicity without the ability to coordinate across persons with a calendar – an invention that stemmed from expanding our spatial capabilities beyond their initial domain.
Language, arguably the deepest propellant behind organized conscious thought, has also benefited from the brain’s spatial processing foundation. I remember that first time I met with the philosopher and neuroscientist Dr. Giulio Tononi. Although his articulate phrasing and obvious mastery of consciousness theory relayed his thoughts to my mindscape in an exceedingly clear fashion, I found myself enthralled by the movement of his hands as he spoke. Dr. Tononi talking with his hands, as is commonplace amongst Italians, is an example of “embodied cognition”. First termed by Rosh et al. (1992), embodied cognition captures the notion that “cognition depends upon the kinds of experience that come from having a body with various sensorimotor capacities, and second, that these individual sensorimotor capacities are themselves embedded in a more encompassing biological, psychological and cultural context” (Rosh, 1999). The dependence of cognition upon the body and its interactions with the environment illustrates that cognition is not an end in itself but is constrained by an organism’s goals and capacities (Shapiro, 2014). However, such constraints do not mean cognition is set by adaptive behavior (or autopoiesis) alone, but cognition requires “some kind of information processing…the transformation or communication of incoming information”, the acquiring of which involves “exploration and modification of the environment”. Milkowski (2013) illustrates this point best:
It would be a mistake, however, to suppose that cognition consists simply of building maximally accurate representations of input information…the gaining of knowledge is a stepping-stone to achieving the more immediate goal of guiding behavior in response to the system’s changing surroundings.
Thus, a framework that was used to represent one’s situatedness[3] in an environment has once again been recruited so as to enhance a separate cognitive domain—language. Gesticulation by way of hand movements has been repeatedly demonstrated to facilitate understanding by way of conveying concepts in a fashion that superposes and extends verbal restraints. Sign language is an explicit example of how the observation of motion can drive thought transfer in an isolated fashion. By creating motion within the domain that recruits our spatial processing mechanisms (i.e. the space between a speaker and a listener) we create a kind of back-channel way of expressing and working out thoughts with optimized machinery.
The thoughts expressed by hand motions are often our newest and most advanced ideas about the problem we’re working on; we can’t yet assimilate these notions into language, but we can capture them in movement
nascent (Goldin-Meadow, 2005).
Cook et al. (2008) solidified this notion of spatially-enhanced-communication by way of research showing that college students who gestured as they retold short stories they’d seen recalled the details of the stories better, suggesting that gesturing as we’re remembering helps to bolster the hoped accomplishments of language. In the conclusions, Cook et al. (2008) go on to say, “gesturing can thus play a causal role in learning, perhaps by giving learners an alternative, embodied way of representing new ideas.” Broaders et al. (2007) provides additional evidence for the eased conveyance of concepts when an instructor utilizes embodied cognition alongside lingual explanations: third-graders who were asked to gesture while learning algebra were nearly three times more likely to remember what they’d learned than classmates who did not gesture. Thus, even language, one of the more integral components of human intelligence, takes a helping hand from the brain’s disposition to represent things within a spatial framework.
If cognition is thought of as an ascending assimilation of information, then reasoning is certainly at its apex. Despite its neural prestige, even analogical reasoning and its associated areas of advanced abstraction, logic, and decision-making have witness benefit from reverse-projecting complex information from the high-dimensional feature space in which it resides into a tangible space realm—the one in which we have perceptual access to. Perhaps the most blatant exemplar of spatial processing recruitment to the domain of reasoning is in the use of spatial limits in illustrating logical possibilities. The notable amelioration of difficult logical problems by way of manipulating them within a spatial framework has its merit based in plausibility space. That is, as a thinker entertains the plausibility of an argument, he can construct a space for each premise to exist in. As subsequent premises are presented, the observer can note their dependence, independence, superposition, etc. with the previous premises. Subsequently, when a conclusion about such relations needs to be evaluated, the thinker can recruit his/her spatial processing framework to identify the spatial plausibility of such a conclusion and then translate that into terms of logical validity. A simple Venn-Diagram accomplishes this concept in an exemplary fashion, as is demonstrated in Figure 1.
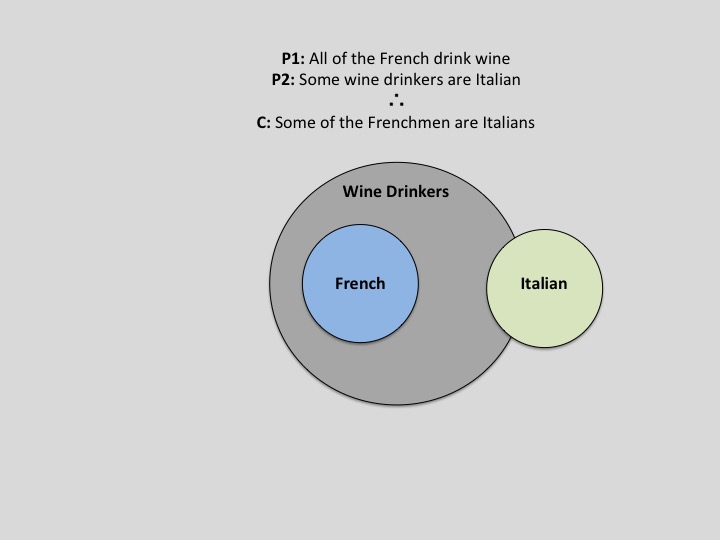
Fig. 1 – An illustration of how Venn-Diagrams can aid in the refutation of logical arguments. Adapted from UCLA lecture notes originally authored by Dr. Keith Holyoak (2015)
While limited in its complexity, the Venn-Diagram serves as an introductory segue to using spatial infrastructures to tackle complex logical problems. A more advanced example would be “Tarski’s World”– a software supplement typically included in introductory courses focused on first-order logic (See Appendix A. for a visualization).
The Tarski’s World application allows the evaluation of first-order sentences within blocks world which users may construct using a simple editor. The worlds consist of collections of blocks of varying sizes and shapes, and placed on a checkerboard. We use an interpreted first-order language, which allows users to write sentences about these worlds and evaluate their truth. A Henkin-Hintikka game may be used to elucidate the evaluation procedure (Barwise, 2993).
Such spatial processing within the reasoning domain seems almost fundamental to a thinker’s advanced success. While the cognitive science community has yet to reach a unanimous consensus on the rightful declaration of an individual’s domain-invariant reasoning abilities, dubbed fluid intelligence[4], many have put faith in the Raven’s Progressive Matrices (Raven, 1936; Raven, 2000) as a test for varying degrees of intelligence. Interestingly enough, Raven’s Progressive Matrices is entirely based on spatial reasoning—identifying a subsequent spatial pattern following a preceding evolution of spatial transformations. Perhaps it is the case that one’s ability to think within a spatial framework speaks directly to their ability to recruit this framework for non-spatial processes and reap its enhancement benefits. Such a scenario sees support from the observation that students who are explicitly exposed to spatial depictions of solutions to analogical problem solving puzzles are more likely to adapt that solution to future problems, especially if they are of higher intelligence (Kubricht, 2015).
Perhaps the most documented across-domain cognitive relationship is between space and memory. Such a relationship has a deep and profound set of explanatory evidence from cognitive neuroscience. The hippocampal formation has long been associated with the encoding of both spatial environments and long-term episodic memories. Researchers have made remarkable progress towards understanding the mechanisms of these seemingly disparate processes. Concerning the encoding of spatial environments, the hippocampal formation has a variety of spatially specialized neurons: “place cells” have been shown to fire specifically at one or a few locations in an animal’s environment (for a review see (O’Keefe, 1979); a population of “border cells” in the entorhinal cortex encode a representation of geometric borders (Solstad et al., 2008); “grid cells” in the entorhinal cortex, which encode the entire expanse of a spatial location via a triangular coordinate system; “head-direction cells” in the postsubiculum discharge as a function of the animal’s head direction in the horizontal plane, independent of the animal’s behavior (Taube et al., 1990). Concerning the encoding of long term episodic memories, the hippocampal formation’s involvement in the establishment of new memories is a frequently observed phenomenon across all domains of neuroscience (Squire and Zola, 1996; Squire, 1992; Tulving and Markowitsch, 1998; Rissman and Wagner, 2012) . Degradation or removal of the hippocampus is strongly correlated with the inability to form new memories (Scoville and Milner, 1957). Additionally, individual differences in hippocampal integrity correlate with mnemonic function (Simoneit and Hites, 1997).
A growing body of literature has attempted to ascribe a common cause as to why the hippocampal formation is important for both memory and space (Bird and Burgess, 2008; Eichenbaum et al., 1999). The medial pallium, whose allocortex forms the hippocampal formation, evolved alongside human navigation into novel terrain (Jacobs, 2003). Thereby, it should come as no surprise that the hippocampus is primarily a spatial mechanisms and that spatial memory is likely our strongest form of memory. Rudimentarily, both spatial and mnemonic representation in the hippocampus necessitates a network of interconnected representations. Both O’Keefe & Nadel’s spatial map theory (1978) and Eichenbaum & Cohen’s relational memory theory (2001) speak to this point. O’Keefe & Nadel posit that populations of place cells can become interconnected and provide an animal with a flexible, allocentric representation of space that, according to [Best and White, 1999]), can then be used for novel route planning. Similarly, Eichenbaum & Cohen proposed that abstract features of an episode (e.g objects, people, etc.) are individual nodes in a network, whose temporal linkage forms a collective representation of an event. This allows for the ability to re-instantiate the same network with the activation of some, but not necessarily all, nodes within it. Research into the existence of “concept cells” within the hippocampus (Quiroga, 2012) , which are sparsely activated by highly-specific domain-invariant (i.e presentation by picture, audio, or text elicits the same neuronal response) concepts (e.g Luke Skywalker), suggest a hierarchical mechanism that integrates overlapping mnemonic networks in the same way that both grid cells and sensory inputs create succinct place cells (Bush et al., 2014) . Thereby, it appears as though hippocampal circuitry is hierarchical in nature and such a framework is crucial for the encoding of both space and events: space is made up of a collection of “places”, and both events and concepts are made up of a collection of “features”.
The interplay between space and memories is further emphasized by the observation that hippocampal representations of both space and concepts vary anatomically depending on the size of the representation. For example, there is a progressive increase in grid scale from dorsal to ventral medial entorhinal cortex (Brun et al., 2008), meaning that larger and larger grids are formed to overlay the special environment. Researchers have been actively attempting to parallel this anatomical discretization of space by examining the sub-regions of the hippocampus in relation to their role in the construction of either abstract versus detailed concepts/events. Primarily, it has been shown that the anterior hippocampal formation is concerned with the encoding of abstract concepts and memories while the posterior hippocampal formation is concerned with the encoding of more detailed forms of memory (Poppenk et al., 2013; Ranganath and Ritchey, 2012).
As emphasized by Milivojevic and Doeller (2013), the study of the hippocampus’ role in both space and memory has been approached, thus far, in isolation. Subsequently, any theories that have attempted to pinpoint the underlying neural mechanisms that can support both processes are inherently post-hoc and potentially flawed. The aim of this current research is to elucidate the bridges that span the gaps between such hippocampal processes from an a priori perspective. In other words, we sought to unveil how the human hippocampus recruits a circuitry that initially evolved for the processing of spatial environments and use it for the memorization of non-spatial content. In so doing, we examine the ways in which space and memory become intertwined and can, through scientific operations, be dichotomized.
Fantastically positioned at this junction of space and memory, lies the most widely used memory enhancement technique: The Method of Loci. The Method of Loci is first noticed in Marcus Cicero’s De Oratore (written in 55BCE – translated by A. S. Wilkins Clarendon in 1963). Cicero in his De Oratore shares the story of the poet Simonides of Ceos who was dining whilst sharing his poetry in Thessaly at a large banquet table. Halfway through his poem, Simonides was called for a message outside. During this time, the building collapsed with such destruction that it not only killed all the guests inside, but left their bodies unidentifiable. Simonides was left with the task of attempting to relay to the deceased’s relatives which body belonged with which name. Simonides was astounded by his ability to recall the locations where each of the guests (whom he had just met that night) had sat. He was subsequently able to parlay this information for the identification of the mangled bodies. As a poet in Ancient Greece, Simonides was previously enthralled with the art of memory; spellbinding oration for the presentation of poetry mandates an intimate acquaintance with the verse at hand. “Noting that it was through his memory of the places at which the guests had been sitting that he had been able to identify the bodies, he realized that orderly arrangement is essential for good memory” (Yates, 1966). Simonides went on to develop a mnemonic method that could bundle the benefits he observed that fateful day.
Over 2000 years later, the instructions that Simonides solidified remain, fundamentally, the same. In order to implement the Method of Loci, one is typically instructed to conjure up a familiar structure (e.g., a childhood home) in their imagination that will serve as their “memory palace”. While mentally navigating through this environment, one can imagine “placing” a list of to-be-remembered items in different locations. When later attempting to retrieve this list of items, one simply has to navigate back through the environment and “observe” the objects in their previously placed locations.
The efficacy of the Method of Loci in enhancing memory is a well-documented behavioral phenomenon (Ross and Lawrence, 1968; Crovitz 1971; Briggs 1971). Subjects often times witness a seven-fold increase in ordered retention (Bower, 1970). Its potential for increasing memory has been used by memory champions and educators to immense success (Foer, 2011). The incredible mnemonic effects witnessed by users of The Method of Loci should come as no surprise. Seeing as the hippocampus primarily evolved as a spatial machine (Jacobs, 2003), it would follow that it’s most impressive feat is that of spatial encoding. Thereby, it follows that the Method of Loci seems to work by exploiting the remarkable vividness of our visuospatial memories by using spatial environments as scaffolding for memorizing non-spatial content.
In conclusion, thinking with space appears to be a practically omnipresent phenomenon throughout all of cognition. While the introduction to this text spoke to the literal impossibility of thinking without space, it appears as though the figurative definition is equally inseparable. Projecting problems into a spatial domain reliably facilitates the solving of that problem by taking advantage of the brain’s spatial processing system, which has an insurmountable temporal advantage of advancement due to its early adoption by organisms in order to accomplish the most basic tasks of survival. Such a postulate has observable neuroanatomical correlates (e.g. hippocampal evolution), a vast array of societal implications (e.g. calendars and analog watches aiding culture) and behavioral benefits (e.g. facilitating communication, reasoning, and bolstering memory strength). As the cognitive and educational sciences begin to incorporate explicit spatial learning aids (e.g. through the use of virtual reality) into research and instruction, it is reasonable to anticipate a revolutionary, all-encompassing enhancement of thought by way of a poetic recycling of the systems that allowed for thought in the first place.
Appendix A.
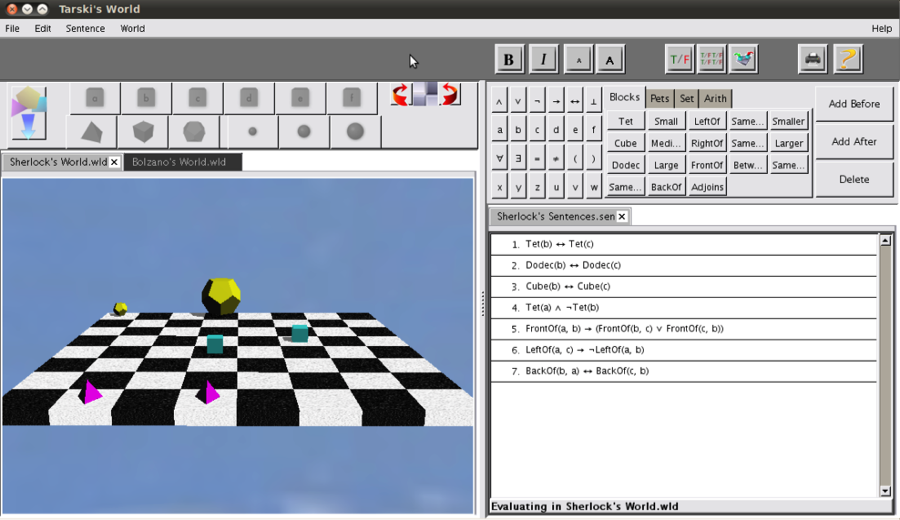
Tarski’s World — software supplement typically included in introductory courses focused on first-order logic. Only one shape can occupy any one place. It can be of only one size and can have potential relations with other objects within the checkerboard
(e.g. a green medium cube can be in front of the small purple pyramid)
Footnotes:
[1] Researchers have since revealed evidence that the tunicate does indeed retain a commendable neural system. However, glial cells are absent and neuronal axons are unsheathed resulting in the functional equivalent of a peripheral nerve net, capable only of generating local activity and mediating simple reflexes (Mackie, 2005). Nonetheless, even in its most primitive form, the example of the sea squirt upholds the subsequent postulation in the above body text.
[2] In addition to calendars, it’s a small leap to recognize the ways in which the analog watch represents time as a sinusoidal harmonic oscillation in its phase-plane—spatial degrees around a circle. Such temporal precision has afforded even more precise instances of synchronicity than the calendar.
[3] In artificial intelligence and cognitive science, the term situated refers to an agent, which is embedded in an environment (Hendricks, 1996). Essentially, for one to be situated in an environment, one must have a sense of self and the space around them– thus making embodied cognition, by way of recruiting neural mechanisms necessary for situatedness, relies upon the brain’s spatial processing mechanisms.
[4] Intelligence as defined by ones capacity for logic, abstract thought, problem solving, creativity, et
References
Babylonian calendars (Encyclopedia Britannica Online) http://www.britannica.com/EBchecked/topic/89368/calendar/60208/Babylonian-calendars
Barwise, Jon, and John Etchemendy. “The Language of First-Order Logic Including the Macintosh Version of Tarski’s World 4.0.” (1993).
Bird, C., and Burgess, N. (2008). The hippocampus and memory: insights from spatial processing. Nature reviews. Neuroscience 9, 182–94
Bower, Gordon H. “Imagery as a relational organizer in associative learning.” Journal of Verbal Learning and Verbal Behavior 9.5 (1970): 529-533
Briggs, Gary G., Stephen Hawkins, and Herbert F. Crovitz. “Bizarre images in artificial memory.” Psychonomic Science 19.6 (1970): 353-354
Broaders, Sara C., et al. “Making children gesture brings out implicit knowledge and leads to learning.” Journal of Experimental Psychology: General 136.4 (2007): 539.
Brun, V., Solstad, T., Kjelstrup, K., Fyhn, M., Witter, M., Moser, E., and Moser, M.-B. (2008). Progressive increase in grid scale from dorsal to ventral medial entorhinal cortex. Hippocampus 18, 1200–12.
Bush, D., Barry, C., and Burgess, N. (2014). What do grid cells contribute to place cell firing? Trends in Neurosciences 37.
Cook, Susan Wagner, Zachary Mitchell, and Susan Goldin-Meadow. “Gesturing makes learning last.” Cognition 106.2 (2008): 1047-1058.
Crovitz, Herbert F. “The capacity of memory loci in artificial memory.” Psychonomic Science 24.4 (1971): 187-188
Dennett, Daniel C. Consciousness explained. Penguin UK, 1993.
Foer, J. (2011). Moonwalking with Einstein: The art and science of remembering everything. New York, NY: Penguin Press
Goldin-Meadow, Susan, and Susan M. Wagner. “How our hands help us learn.”Trends in cognitive sciences 9.5 (2005): 234-241.
Hand Gestures Say A Lot About Your Intelligence (Business Insider)By: Paul, Annie.
http://www.businessinsider.com/hand-gestures-reveal-complex-thoughts-2013-6
Hendriks-Jansen, Horst. Catching ourselves in the act: Situated activity, interactive emergence, evolution, and human thought. MIT Press, 1996.
Jacobs, L. (2003). The Evolution of the Cognitive Map. Brain, Behavior and Evolution 62, 128–139
Johnson, Brian, and Ben Shneiderman. “Tree-maps: A space-filling approach to the visualization of hierarchical information structures.” Visualization, 1991. Visualization’91, Proceedings., IEEE Conference on. IEEE, 1991.
Kemp, Barry J. Ancient Egypt: anatomy of a civilization. Psychology Press, 2006.
Kubricht, James, Lu, Hongjing, Holyoak, Keith. In Press. “Animation Facilitates Source Understanding and Spontaneous Analogical Transfer”. Proceedings of the 37th Annual Conference of the Cognitive Science Society.
Llinás, Rodolfo. “I of the Vortex. From Neurons to Self. 2001.” (2001).
Mackie, G. O., and P. Burighel. “The nervous system in adult tunicates: current research directions.” Canadian journal of zoology 83.1 (2005): 151-183.
Milivojevic, B., and Doeller, C. F. (2013). Mnemonic Networks in the Hippocampal Formation: From Spatial Maps to Temporal and Conceptual Codes. Journal of experimental psychology. General. Available at: http://www.ncbi.nlm.nih.gov/pubmed/23875564
Miłkowski, Marcin. Explaining the computational mind. Mit Press, 2013.
O’Keefe, J. (1979). A review of the hippocampal place cells. Progress in Neurobiology 13, 419439.
O’Keefe, J., & Nadel, L. (1978). The hippocampus as a cognitive map. Oxford, England: Oxford University Press.
Poppenk, J., Evensmoen, H., Moscovitch, M., and Nadel, L. (2013). Long-axis specialization of the human hippocampus. Trends in cognitive sciences 17, 230–40.
Quiroga, R. Q. (2012). Concept cells: the building blocks of declarative memory functions. Nature reviews. Neuroscience 13, 587–97.
Ranganath, C., and Ritchey, M. (2012). Two cortical systems for memory-guided behaviour. Nature reviews. Neuroscience 13, 713–26.
Raven, J. C. “Mental tests used in genetic studies: The performance of related individuals on tests mainly educative and mainly reproductive.” Unpublished master’s thesis, University of London (1936).
Raven, John. “The Raven’s progressive matrices: change and stability over culture and time.” Cognitive psychology 41.1 (2000): 1-48.
Rissman, J., and Wagner, A. (2012). Distributed representations in memory: insights from functional brain imaging. Annual review of psychology 63, 101–28.
Rosch, Eleanor, Evan Thompson, and Francisco J. Varela. The embodied mind: Cognitive science and human experience. MIT press, 1992.
Ross, John, and Kerry A. Lawrence. “Some observations on memory artifice.” Psychonomic Science (1968)
Scoville, W., and Milner, B. (1957). LOSS OF RECENT MEMORY AFTER BILATERAL HIPPOCAMPAL LESIONS. Journal of Neurology, Neurosurgery & Psychiatry 20, 11–21.
Shapiro, Lawrence, ed. The Routledge handbook of embodied cognition. Routledge, 2014.
Simoneit, B., and Hites, R. (1997). Differential Effects of Early Hippocampal Pathology on Episodic and Semantic Memory. Science 277, 376380
Solstad, T., Boccara, C., Kropff, E., Moser, M.-B., and Moser, E. (2008). Representation of Geometric Borders in the Entorhinal Cortex. Science 322, 1865–1868.
Squire, L. R. (1992). Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psychological review 99, 195–231.
Squire, L. R., and Zola, S. M. (1996). Structure and function of declarative and nondeclarative memory systems. Proceedings of the National Academy of Sciences 93, 13515–13522.
Talk With Your Hands: Gestures Are Important (Ideas The Secret Code Of Learning Comments)
Taube, J. S., Muller, R. U., and Ranck, J. B. (1990). Head-direction cells recorded from the postsubiculum in freely moving rats. I. Description and quantitative analysis. The Journal of neuroscience : the official journal of the Society for Neuroscience 10, 420–35.
Tulving, E., and Markowitsch, H. (1998). Episodic and declarative memory: Role of the hippocampus. Hippocampus 8, 198–204
Wallace, David Foster. This is water: Some thoughts, delivered on a significant occasion, about living a compassionate life. Hachette UK, 2009.
Wolpert, Daniel. (2011, July). The real reasons for brain. Retrieved from http://www.ted.com/talks/daniel_wolpert_the_real_reason_for_brains?language=en
Yates, Frances Amelia. The art of memory. Vol. 64. Random House, 1992.

This is very interesting. I am drawn to it because of my interest in how the mind can subconsciously use spatial relationships to organize data. Are you interested in that as well?
I am most certainly interested in that! I tend to think more about the volitional ways in which we organize data spatially. However, the subconscious component would also be quite interesting– although admittedly difficult to formally study. Any ideas on how to approach a scientific design aimed at elucidating the subconscious mechanisms at play here?
yes thinking with/in/through space, mental space, is our, human way of representing anything.
Animals have and need no “extra” space, their Umwelt works through direct, immediate perception, without (a notion of) our (paradox: invisible yet material/airy “medium”) space.
Air and light are not known to animals, therefore animals are not separated from things or each other.
…says fuerlinger@hominisation.at